EL METABOLISMO DE HIDRATOS DE CARBONO
El metabolismo de hidratos de carbono es
una de las principales rutas de metabolismo celular .entre los azucares
utilizados como gran fuente de energía para la célula destaca principalmente la
glucosa ya que es la base de muchos polisacáridos la glucosa ocupa un papel muy
funcional y central en el metabolismo de los hidratos de carbono
También conoceremos las principales rutas
metabólicas presentes en el organismo que se van a relacionar estrechamente con
la producción de energía y de poder reductor ya que la energía que se utiliza
generalmente proviene del metabolismo de los azucares como la glucosa.
Las rutas relacionadas con el glucógeno
polisacárido de reserva energética de síntesis más rápida o a corto plazo es la
de los animales. Es importante conservar un buen estatus energético en el
organismo especialmente en músculos e hígado
La glogenolisis comprende la reacción de
degradación mientras que la gluconeogénesis incluye la vía de síntesis a partir
de la glucosa.
De las ruta relacionada con los
monosacáridos tenemos como principal la
ruta catabólica llamada glucolisis ruta degradativa de la glucosa que sirve para la obtener
energía de esta molécula y también de otras hexosas y monosacáridos
La glucogénesis es la principal ruta
anabólica que sintetiza glucosa a partir de intermediarios metabólicos como
intermediario metabólico tendremos al piruvato que participa como vías de
entrada en las rutas catabólicas de fermentaciones
El análisis
y las comprensión de este conjunto de rutas complementara el estudio digestivo de los hidratos de carbono ya que
en el caso de los humanos animales y
organismos organotrofos, la indigesta de azucares, polisacáridos constituye principalmente
la fuente de hidratos de carbono. Aunque esto organismos pueden sintetizar
azucares a partir de moléculas intermediarias.
LA DIGESTIÓN DE LOS AZUCARES EN LA
DIETA
Todos los organismos superiores cuentan
con un sistema especializado para realizar la digestión de los diferentes
componentes de los alimentos. En el caso de los carbohidratos, el problema es
relativamente sencillo, puesto que la variedad de las moléculas que se ingieren
no es muy grande. En su mayor parte estas son almidones, dextrinas (y
glucógeno en pequeña proporción), sacarosa y galactosa. Desde el punto de vista
de la digestión el problema de los organismos se reduce a convertir estas
moléculas en los monosacáridos que las componen.
En teoría, en los animales superiores la
digestión del almidón se inicia en la boca. La saliva, principalmente aquella
producida por la parótida, contiene una enzima, la amilasa salival, llamada también ptialina. Esta es capaz de actuar sobre los almidones y sobre el
glucógeno, rompiendo los enlaces alfa-1,4 de tal forma que se separan de dos en
dos los fragmentos de la molécula polimérica. Las moléculas que resultan
entonces, son del disacárido maltosa. Pero la acción de la amilasa salival es
de corta duración; el bolo alimenticio permanece en la boca durante el tiempo
de la masticación y luego es deglutido. En el estómago, el HCl del jugo
gástrico le confiere un carácter ácido, con un pH cercano a 2. El pH óptimo de
la amilasa salival se encuentra cercano a 7, por lo cual una vez llegado el
bolo alimenticio al estómago se suspende la acción de la enzima. Lógicamente el
efecto logrado por ésta no es muy importante, de manera que desde el punto de
vista práctico la digestión de los carbohidratos por la acción de la saliva es
nula. En realidad, la digestión de los almidones se inicia en el intestino
delgado por la acción de la amilasa pancreática, enzima que tiene el mismo
mecanismo que la amilasa salival, es decir que el almidón se va convirtiendo en
maltosa. Además hay otra enzima, la amilo-1,6-glucosidasa, que se encarga de
romper los enlaces alfa-1,6 de manera que la acción combinada de ésta y de la
amilasa da como resultado la conversión total del almidón en moléculas de
maltosa. Luego la maltosa es objeto de la acción de diferentes tipos de
maltasas, producidas por el intestino, que realizan la degradación completa de
la molécula para convertirla exclusivamente en glucosa.
Para la digestión de los disacáridos hay
sendas enzimas producidas por la mucosa intestinal, que los reducen a sus
componentes: la sacarasa convierte a la sacarosa en glucosa y fructosa; la
lactasa convierte a la lactosa en glucosa y galactosa. El final del proceso
digestivo es una mezcla de glucosa, galactosa y fructosa en la cual pre-domina,
desde luego la primera. En la figura 9.1 se presenta en forma esquemática el
proceso digestivo de los carbohidratos. En los humanos se pueden presentar
alteraciones congénitas en la síntesis de algunas de las enzimas digestivas de
los carbohidratos. En algunos casos falta la saca- rasa; con mucha mayor
frecuencia ocurren casos de ausencia de la lactasa, que desde luego es mucho
más seria que la de la primera. Sin embargo, hay que señalar que son raros los
casos de estas deficiencias enzimáticas.
Boca: Amilasa salival: Almidón — Maltosa
Páncreas: Amilasa pancreática: Almidón —
Maltosa
Intestino: Maltasas: Maltosa — Glucosa
Sacarasa: Sacarosa — Glucosa + Fructosa
Lactasa: Lactosa — Glucosa + Galactosa
Una vez digeridos los carbohidratos, el
intestino tiene que introducir las moléculas resultantes en el organismo. Con
este objetivo, dispone de sistemas de transporte específicos para los azúcares,
que además requieren de la participación de los sistemas energéticos para
funcionar. No está claro aún cuál es el mecanismo mole-cular por el que los
azúcares cruzan la pared intestinal. Se sabe que el sistema de transporte es
específico y depende de una fuente de energía, la cual a su vez tiene su origen
en el metabolismo de las células de la pared intestinal, y finalmente, pare-ce
haber una cierta relación entre el transporte de los azúcares y el del sodio.
De cualquier manera, el hecho es que los monosacáridos resultantes de la
digestión de los carbohidratos son absorbidos por la pared intestinal, y
aparecen luego en el torrente circulatorio de la vena porta, que los lleva a
todo el organismo, pasando primero por el hígado, para su distribución general.
La glucólisis es una vía que permite
obtener ATP a las células.
La glucólisis (o glicólisis) es una vía
catabólica a través de la cual tanto las células de los animales como
vegetales, hongos y bacterias oxidan diferentes moléculas de glúcidos y
obtienen energía. El hecho de que esta vía ocurra en organismos muy diversos,
indica que es una vía metabólica conservada, es decir presente en organismos
filogenéticamente distantes.
•Para su estudio, describiremos 9 reacciones
enzimáticas que ocurren en el citoplasma y permiten la transformación de una
molécula de glucosa a dos moléculas de piruvato. La degradación hasta piruvato es parte del
proceso catabólico o degradativo de los glúcidos, porque estas moléculas pueden
seguir oxidándose y continuar entregando energía a la célula.
Esquema
de la Glucólisis. Se representan los principales intermediarios, su número de
carbonos (C) y las fases de consumo y producción de ATP (primera y segunda fase
respectivamente)
El balance neto para la reacción global de
la glucólisis es:
Hexosa + 2 NAD+ + 2 ADP + 2 Pi 2 NADH + 2 piruvato + 2 ATP
En la glucólisis se pueden establecer dos
fases
Primera fase = Activación de la hexosa
(glucosa por ej.), con gasto de energía como ATP.
Segunda fase = Obtención de energía que se
conserva como ATP.
• La primera fase es endergónica, porque
se consumen 2 ATP, y consta en la transformación de una hexosa (por ejemplo,
glucosa) en dos triosas (dihidroxicetona 3 P y gliceraldehído 3P). La segunda
fase es exergónica, dado que se forman 4 ATP utilizando la energía liberada de
la conversión de 2 gliceraldehídos 3P en 2 piruvatos .
• La glucólisis ocurre a través de
reacciones enzimáticas, donde cada enzima cataliza una reacción o paso
específico. De esta forma, cuando se hace referencia a una isomerasa, lo es a
una específica para determinada molécula, y no a una isomerasa universal que
catalice cualquier reacción de isomerización. Lo mismo sucede con las quinasas,
deshidrogenasas, etc.
Visualizar
el conjunto de reacciones que conforma a la glucólisis, previo a la descripción
de cada reacción, ayuda a tener una idea general sobre lo que incluye esta vía,
que transcurre en el citoplasma. En la figura 2 se observa, al igual que en la
figura 1, la etapa de inversión de energía y la de síntesis de ATP, así como a
partir de una hexosa, en este caso la Glucosa, se obtienen dos moléculas de
piruvato, de 3C cada una.
Encontramos un esquema general con la
secuencia de reacciones que incluye la glucólisis. GA3P, gliceraldehído 3-P;
D3P, dihidroxicetona 3-P. Se numeran las
reacciones tal cual están descriptas en el texto.
La
fase de gasto de energia va desde una hexosa no fosforilada hasta el GA3P y
D3P.
La
glucosa, se fosforila y rinde glucosa 6P (G6P), una molécula con mayor energía.
La enzima responsable de la reacción, una quinasa (hexoquinasa) consume una
molécula de ATP y libera ADP. La misma hexoquinasa fosforila otras hexosas como
fructuosa, galactosa y manosa.
Es irreversible, es decir la los
productos (G6P y ADP) no liberan los reactivos (Glucosa y ATP).
La fosforilación de la glucosa tiene
ventajas para la célula: la G6P es más reactiva que la glucosa y a diferencia
de ésta no atraviesa la membrana celular porque no tiene transportador. De esta
forma se evita la pérdida de un sustrato energético para la célula.
La
G6P se isomerisa a fructosa-6-fosfato (F6P) por acción de una isomerasa, que
facilita la isomerización de estas hexosas en los dos sentidos: de F6P a G6P o
de F6P a G6P, la reacción es reversible.
Consiste
en la fosforilación de la F6P en el C1, que rinde fructosa 1,6-bifosfato (F1-6P).
En esta reacción, catalizada por otra quinasa, la fosfofructoquinasa (FFQ), se
consume ATP. Esta enzima merece especial atención porque, como se mencionará
más adelante, participa en la regulación de la glucólisis.
Esta
reacción, al igual que la primera, es irreversible, y ambas constituyen pasos
importantes porque son los puntos de control de la glucólisis.
En
esta reacción la F1-6P se rompe en 2 moléculas de 3 carbonos (triosas): la
dihidroxiacetona 3-fosfato (D3P) y gliceraldehído 3-fosfato (GA3P) mediante una
reacción reversible catalizada por una liasa (aldolasa).
El
GA3P sigue los pasos de la glucólisis, la otra triosa generada, D3P, por
isomerización produce otra molécula de GA3P. La reacción es reversible, y está
catalizada por una isomerasa.
Éste es el último paso de la Fase con
gasto de energía en la que se consumieron 2 ATP.
Así, en el cuarto paso se genera una
molécula de GA3P, y en el quinto paso se genera la segunda molécula de éste. De
aquí en adelante, las reacciones ocurrirán dos veces, debido a que se generan
dos moléculas de GA3P por hexosa.
Hasta el momento solo se han
consumido 2ATP, sin embargo, en la segunda etapa, el GA3P se transforma en una
molécula de alta energía, a partir de la cual se obtendrá el beneficio final de
4 moléculas de ATP.
Consiste en la oxidación del GA3P e
incorporación de un fosfato a la molécula, de manera que se genera un compuesto
con mayor energía. En este paso, que en realidad implica dos reacciones, actúa
una deshidrogenasa que utiliza NAD+ y se genera NADH.H. Se verá al finalizar la
descripción de la vía, cómo y por qué es necesario reoxidar este cofactor.
En
este paso el grupo fosfato del 1,3-bifosfoglicerato se transfiere a una
molécula de ADP, por una quinasa, generando así la primera molécula de ATP de
la vía. Esta manera de obtener ATP, en la que no participa la cadena
respiratoria, se denomina fosforilación a nivel de sustrato.
Como
la glucosa se transformó en 2 moléculas de GA3P se sintetizan un total de 2 ATP
en este paso.
Las
reacciones 6 y 7 de la glucólisis corresponden a un caso de acoplamiento, donde
una reacción energéticamente desfavorable (6) es seguida por una reacción muy
favorable energéticamente (7) que induce a que ocurra la primera (figura 2).
Consideramos
aquí a dos reacciones sucesivas, de las cuales una, la isomerización del
3-fosfoglicerato a 2-fosfoglicerato, no aparece representada en la figura 2 y
la otra corresponde a la transformación del 2- fosfoglicerato en fosfoenolpiruvato
(PEP), por acción de la enolasa.
2-Fosfoglicerato
- PEP +
H2O
En
la última reacción, irreversible, se desfosforila el PEP y se obtiene piruvato
y ATP. La transferencia del grupo fosfato del PEP al ADP la cataliza una
quinasa (piruvato quinasa). Es la segunda fosforilación a nivel de sustrato: se
fosforila el ADP a ATP independientemente de la cadena respiratoria.
Como
se observa, el oxígeno no es necesario en ninguna reacción de la glucólisis; la
vía ocurre en células aerobias y fermentativas.
En la ruta glucolítica existen tres puntos
importantes de control .El primero es aquel en que la glucosa se fosforila a
glucosa 6-fosfato por ATP y la hexoquinasa. Otro importante punto de control es
la reacción catalizada por la fosfofructoquinasa. Esta enzima reguladora es
activada por AMP y ADP e inhibida por ATP y citrato. El tercer punto de
regulación es la reacción catalizada por la Piruvato quinasa, que es activada
por fructosa 1,6-bisfosfato y AME Como se puede observar, las tres enzimas de
control están reguladas por un intermediario metabólico, pero de una manera
especial por la concentración de AME ADP y ATE. De una manera simplificada se
puede afirmar que la relación ADP/ATP regula el flujo metabólico de la ruta. Si
esta relación es alta porque la concentración de ATP es baja, la glucólisis se
activa para formar ATP. Si, por el contrario, la relación ADP/ATP es baja, la
célula inhibe la fosfofructoquinasa y se interrumpe la glucólisis para no
producir más ATP
La hexoquinasa
Es inhibida por el producto de la
reacción, la G-6-P y activada por Pi. La isoenzima de la hexoquinasa en hígado
se llama glucoquinasa y tiene menor afinidad por la glucosa que la HK, luego
tendrá una KM más alta.
La
fosfofructoquinasa 1 (PFK1) Es la enzima clave en el
control de la glucolisis; es una enzima alostérica y está regulada por
metabolitos activadores (F-2,6-BP, AMP) y otros inhibidores (ATP, citrato, H+
).
La
Piruvato quinasa
Es
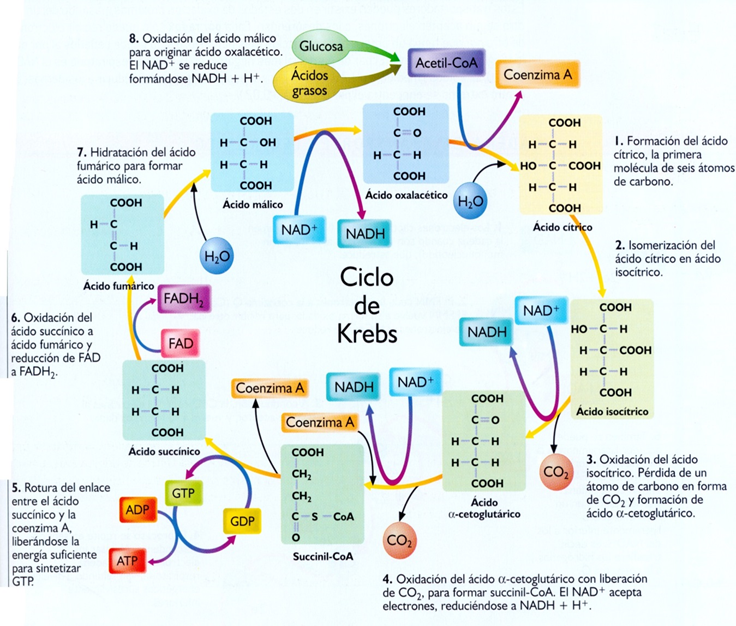
inhibida por el ATP, ALA, Acetil-CoA y los ácidos grasos de cadena larga. Los
últimos pueden proporcionar ATP a través del Ciclo de Krebs. Es activada por
F1,6-BP. En hígado es inhibida por fosforilación. Metabolismo de
Fructosa-2,6-bisfosfato
La fructosa-2,6-bisfosfato es un activador
alostérico de la PFK1, el que la activa más potentemente. La fructosa-2,6-bisfosfato
se forma desde la fructosa-6-fosfato en reacción catalizada por la
fosfofructoquinasa 2 (PFK2). La F-2,6-BP es activador alostérico de la PFK-1,
siempre que exista AMP. Es decir, para anular la inhibición del ATP, el AMP y
la F2,6-BP deben estar presentes. La fructosa-2,6-bis-P impide que el flujo
glicolítico se detenga cuando haya ciertos niveles de ATP en la célula. La PFK2
es una actividad que radica en una proteina bifuncional (proteína con dos
funciones enzimáticas) junto con la actividad F-2,6-Bisfosfatasa, por tanto
puede catalizar la síntesis y la degradación de la F-2,6-BP, según esté
fosforilada o sin fosforilar. Estos razonamientos se completarán en el estudio
de la gluconeogénesis
Incorporación de otros glúcidos a la
glucolisis o Rutas alimentadoras de la glucolisis Polisacáridos (Glucógeno,
almidón).- El primero se degrada por la vía de la glucogenolisis hasta unidades
de glucosa-1-P, que se incorporan a la fase preparatoria de la glucolisis para
su degradación.
Disacáridos (disacaridasas).- Su
hidrólisis produce monosacáridos que se incorporan a la glucolisis por
diferentes vías Sacarosa + H2O -------------> fructosa + glucosa sacarasa
Lactosa + H2O --------------> galactosa + glucosa lactasa Maltosa + H2O
--------------> 2 glucosa maltasa.
Para evitar los pasos irreversibles que se
originan en la glucólisis. la gluconeogénesis utiliza una serie de reacciones
alternativas catalizadas por enzima:. Diferentes. En la figura se muestra un
esquema de la ruta gluconeogenica, enfrentada a la ruta glucolitica. Los tres
pasos irreversibles de la glucólisis se solventan a través de las siguientes
reacciona, que son termodinámicamente favorables:
1.
Síntesis de fosfoenolpiruvato: la conversión del piruvato en fosfoenolpi-ruvato
requiere dos reacciona catalizadas por sendas enzimas: la piruvato carboxilasa,
que canina la conversión de piruvato en oxalacetann y la fosfoenolpiruvato
carboxiquinasa que cataliza la conversión del oxalacetato en fosfoenolpiruvato.
La primen enzima, la Piruvato
carboxilasa, requiere el gasto de una molécula de ATP para fijar un
nuevo átomo de carbono, procedente del CO, para generar oxalacetato, proceso
que exige biorina como cofactor enzimitico. La conversión de una molécula de
tres carbonos en otra de cuatro ocurre en la mitocondria. Posteriormente la
hidrólisis del GTP impulsa la transformación del ~lacean, en fos-foenolpiruvato
y CO2. gracias a la fosfoenolpiruvato carboxiquinasa, en-zima que puede actuar
canco en la mitocondria como en el citoplasma celular dependiendo de la
especie. Posteriormente, el fosfoenolpiruvato se convertirá en
fructosa-1.6-bifosfato siguiendo, en sentido contrario, las reacciones
reversibles de la glucólisis, ya descritas. 2. Conversión de la fructosa,
6-bifosfato en fructosa-6•fosfato: ésta es una reacción hidrolítica por la cual
se elimina el grupo fosfato en posición 1 de la fructosa por acción de la
enzima fructosa-1,6-bifosfatasa. En esta reacción no se regenera ATP si no que
se obtiene P. La fructosa-6-fosfato se convertirá en glucosa-6-fosfato por la
reacción reversible detallada en el apartado de la glucólisis.

Regulación
de la gluconeogénesis
Es la síntesis de glucosa a partir de
precursores no carbohidratos, es decir, síntesis de azúcares a partir de no
azúcares. Los principales precursores son lactato, aminoácidos, oxalacetato y
glicerol.
No se puede formar glucosa a partir de
ácidos grasos porque se convierten en Acetil-CoA y los carbonos se pierden en
forma de CO2.
Ocurre en animales, plantas, hongos y
microorganismos; es una ruta universal. La gluconeogénesis es la principal
fuente de glucosa en ayuno para evitar un shock hipoglucémico. En humanos esta
ruta tiene lugar mayoritariamente en el hígado pero también en los riñones.
Comparte reacciones con la glucólisis con la excepción de las catalizadas por
HK, PFK y PK (reacciones de no equilibrio). Como todo proceso anabólico
requiere un aporte de energía.
Conversión de piruvato en fosfoenolpiruvato:
piruvato carboxilasa & PEPCK
Necesitamos dos enzimas para pasar de
piruvato a fosfoenolpiruvato (2 de las reacciones anapleróticas que reponen los
intermediarios del ciclo de Krebs), estas dos reacciones son exergónicas:
̶
Fosfoenolpiruvatocarboxilasa (necesita biotina). Ocurre en la
mitocondria.
̶
Fosfoenolpiruvatocarboxiquinasa (PEPCK). Ocurre en el citosol.
¡RECUERDA!
Para formar glucosa me hacen falta 2 piruvatos.
>
Rutas alternativas desde el piruvato a fosfoenolpiruvato.
En función del precursor gluconeogénico
(lactato o piruvato) predomina una ruta u otra. La importancia de las rutas
está determinada por la disponibilidad de lactato y las necesidades citosólicas
de NADH en la gluconeogénesis.
El PEP sale de la mitocondria a través de
transportadores.
>
Balance de la conversión de piruvato en PEP:
Piruvato +
ATP + GTP
" PEP +
ADP + GDP
+ Pi
Conversión
de fructosa 1,6-bifosfato en fructosa 6P: fructosa bifosfatasa
La FBPasa-1 promueve la hidrólisis
prácticamente irreversible del fosfato en C-1, en estos casos no la
transferencia de grupo fosforilo al ADP.
Fructosa 1,6-bifosfato + H2O "
fructosa 6P + Pi
Conversión
de glucosa 6P en glucosa: glucosa 6-fosfatasa
Se necesitan 5 proteínas para transformar
la G6P citosólica en glucosa libre. Varias proteínas del RE (retículo
endoplasmático) juegan su papel en la generación de glucosa; T1 transporta la
G6P al lumen del RE, mientras que T2 y T3 transportan Pi y glucosa de vuelta al
citoplasma. La G6P se estabiliza gracias a una proteína que une Ca2+.
2Piruvato + 2ATP + 2CO2 + 2GTP + 2H2O +
2ATP + 2NADH + 2H+ + 2H2O
Glucosa + 2ADP + 2Pi + 2GDP + 2CO2 + 2ADP
+ 2Pi + 2NAD+ + 2Pi
2Piruvato + 4ATP + 2GTP + 4H2O + 2NADH +
2H+ " Glucosa + 4ADP + 2GDP + 6Pi +
2NAD+
La gluconeogénesis no es el proceso
inverso de la glucólisis ya que la célula tiene que gastar 6 enlaces fosfato
ricos en energía, por lo tanto es caro energéticamente fabricar glucosa a
partir de compuestos no carbohidratados.
En los seres humanos los órganos
gluconeogénicos son el hígado y los riñones. Las células inmunes producen
lactato, aspartato y glutamato que van al hígado para la gluconeogénesis.
También interviene el tejido adiposo que produce glicerol mediante degradación
de grasas.
 El glicerol proviene de la hidrólisis de
triacilglicéridos de los adipocitos que llegan al hígado dónde se transforma en
glucosa mediante la siguiente vía:
El glicerol proviene de la hidrólisis de
triacilglicéridos de los adipocitos que llegan al hígado dónde se transforma en
glucosa mediante la siguiente vía:
Gluconeogénesis
desde alanina y otros aa:
Sistema
de transporte del nitrógeno desde el músculo hasta el hígado. La alanina
proviene de la transaminación del piruvato con glutamato lo que forma alanina y
α-cetoglutarato. Cuando la alanina llega al hígado participa en una reacción
semejante pero en sentido contrario produciendo piruvato y glutamato. El
piruvato ya puede entrar en la ruta gluconeogénica
Durante un sprint las células musculares
transforman el glucógeno en glucosa y la consumen inmediatamente, produciendo
piruvato y lactato (porque no hay O2 suficiente). El lactato también va al
hígado dónde se transforma en piruvato para transformarse en glucosa como la
hace el glicerol, glucógeno y aa.
El músculo cardiaco necesita un aporte
continuo de glucosa y la consume siempre en condiciones aerobias.
En ausencia de O2, y en algunas células, el piruvato se
metaboliza hacia compuestos más reducidos para recuperar el NAD+, necesario
para que siga actuando la vía glucolíticas; manteniendo así constante la
relación NAD+/NADH citoplasmática.
La fermentación es la degradación de glucosa en ausencia de
oxígeno; comprende las reacciones glucolíticas y otras reacciones de reducción
finales. Algunos microorganismos y las células musculares, en anaerobiosis,
reducen el piruvato a lactato. Así pueden regenerar el NAD+ necesario para
continuar la glucolisis.
Los microorganismos fermentativos transforman el piruvato
hasta etanol, en dos reacciones: descarboxilación y reducción. Ésta última
permite a las células recuperar el NAD+ necesario para la glucolisis.
Las células que metabolizan en condiciones aeróbicas no
necesitan reducir el NADH para poder continuar la glucolisis, puesto que este
coenzima reducido, NADH, descarga los e- en la cadena respiratoria mitocondrial
y en consecuencia, las células pueden disponer fácilmente de NAD+ para
continuar la glucolisis. En estas condiciones, el piruvato entra en las
mitocondrias, donde se descarboxila y oxida hasta Acetil-CoA. La reacción la
cataliza el complejo enzimático llamado piruvato deshidrogenasa (PDH).
COMPLEJO DE
LA PIRUVATO DESHIDROGENASA
Está
formado por tres actividades enzimáticas y su actuación necesita de cinco
cofactores.
La
actividad de la PDH está regulada alostéricamente por varios metabolitos, como
se aprecia en la siguiente figura. Los metabolitos que indican alta energía
celular la inhiben y el AMP la activa.
Además,
en células eucarióticas, la PDH esta regulada por fosforilacion /
defosforilacion en respuesta a la acción hormonal. La enzima E1 se inactiva por
fosforilación en un resto de SER.
La ruta de las pentosas fosfato,
del fosfogluconato o de las hexosas fosfato ocurre en el citoplasma. Es fuente
de NADPH y ribosa5P para biosíntesis de ácidos nucleicos. Tiene
una fase oxidativa (generación de NADPH) y otra no oxidativa (interconversión
no oxidativa de azúcares).
Fase
oxidativa de la RPF.
Se pasa de una hexosa a una
pentosa. Consiste en dos oxidaciones que convierten la glucosa 6P en ribulosa
5P y reducen el NADP+ a NADPH.
þ La
principal y primera enzima es la G6PDH o G6PD (glucosa 6P
deshidrogenasa).
þ La
segunda enzima es la fosfoglucolactonasa que forma el
6-fosfogluconato que es un metabolito exclusivo de esta ruta y por eso da
nombre a la misma.
þ La
tercera enzima y última de esta fase es la 6-fosfogluconato deshidrogenasa que
forma la ribulosa5P.
>
Balance de la fase oxidativa.
Glucosa6P + 2NADP+ + H2O
"
Ribulosa5P + 2NADPH + 2H+ + CO2
> Fase
no oxidativa.
Comprende pasos que convierten
pentosas fosfato en glucosa6P, la cual inicia el ciclo de nuevo. En esta rama existen
4 tipos de reacciones:
Ü Fosfopentosa
isomerasa: convierte una cetosa (ribulosa) en aldosa
(ribosa).
Ü Fosfopentosa
epimerasa: epimeriza[1] el C3
(convierte ribulosa en xilulosa).
Ü Transaldolasa: transfiere
unidades de 3C.
Ü Transcetolasa:
transfiere unidades de 2C.
En esta fase se utilizan 3
ribulosas5P para formar 2 fructosas6P y un gliceroaldehido3P.
Todas las reacciones de la rama
no oxidativa pueden ocurrir en sentido contrario ya que son próximas al
equilibrio.
> Balance de la fase no
oxidativa.
3Ribulosa5P (3x5C) "
2Fructosa6P (2x6C) + gliceroaldehido3P (3C)
> Balance global.
3Glucosa6P + 6NADP+ +
3H2O "
3Ribulosa5P + 6NADPH + 6H+ + 3CO2
> Relación
de la ruta PPP con otros procesos metabólicos.
Procesos que requieren
NADPH:
· Síntesis
·
Detoxificación
> Regulación
de la ruta PPP.
La entrada de glucosa6P en la
ruta de las pentosas fosfato (RPF) es controlada por la concentración celular
de NADPH que es un fuerte inhibidor de la G6PDH. Como el NADPH es utilizado en
varias rutas metabólicas la inhibición se suaviza y la enzima se acelera para
producir más NADPH.
La síntesis de G6PDH se induce
por el incremento de la ratio insulina/glucagón después de una comida con alto
contenido en carbohidratos. Esto es fundamental para la síntesis de ácidos
grasos
El glucógeno es un polisacárido de origen
animal, formado por una gran cantidad de moléculas de glucosa presenta una
estructura ramificada. La función de este polisacárido es constituir un
reservorio de moléculas de glucosa con la finalidad de cubrir la necesidad a
corto plazo de este monosacárido; por lo tanto el glucógeno sirve como una
reserva de energía a corto plazo. Aunque todas las células pueden tener
glucógeno, este principalmente se forma en dos tejidos: el musculo y el hígado.
El tejido muscular va a utilizar las reservas de glucógeno para cubrir las
necesidades propias de este tejido, especialmente en los momentos de un
ejercicio intenso, mientras que el hígado almacena el glucógeno con la
finalidad de mantener los niveles de glucosa en sangre; en este sentido, el
hígado también se ayuda de la gluconeogénesis para sintetizar glucosa y
mantener los niveles de sangre.
Mantener los niveles de glucosa dentro de
unos límites aceptables es importante, ya que es la glucosa disponible en
sangre la que utiliza la mayoría de los tejidos para obtener la energía que
necesitan en circunstancias normales. Los niveles de glucosa en sangre son
cruciales para una serie de células (entre las cuales se encuentran los
eritrocitos) que obtienen toda su energía de esta glucosa circulante, puesto
que no pueden aprovechar otras fuentes de energía como pudieran ser los ácidos
grasos y los cuerpos cetónicos. Las células cerebrales también dependen en gran
medida de la glucosa en sangre, aunque estas si pueden adaptarse y obtener
energía de los cuerpos cetónicos.
El metabolismo del glucógeno se puede dividir en
dos procesos: la glucogenogénesis o
síntesis de glucógeno y la glucogenolisis
o degradación del glucógeno. Estos son dos procesos opuestos: el primero es
anabólico mientras que el segundo es catabólico.
GLUCOGENOGÉNESIS
La síntesis de glucógeno se produce
normalmente después de la ingestión, sobre todo si la dieta es rica en carbohidratos,
pues en esos momentos habrá una gran cantidad de glucosa en sangre precedente
de la dieta. Esta glucosa será almacenada tanto en el tejido muscular como el
tejido hepático en forma de glucógeno. El tejido hepático será el que se
encargue de acumular la mayor cantidad de glucosa en forma de glucógeno, debido
a
la presencia de la glucoquinasa, enzima
que permite almacenar gran cantidad de glucosa en la célula en forma de
glucosa-6-fosfato. Al contrario que la hexoquinasa presente en los demás tejidos,
esta enzima hepática no se inhibe por la glucosa-6-fosfato, lo cual permite
introducir una mayor cantidad de glucosa dentro de las células.
La glucogenogénesis comienza con la
transformación de la glucosa-6-fosfato en glucosa-1-fosfato por acción de la
fosfoglucomutasa, a través de una reacción reversible. Posteriormente se va a
trasformar en UDP-glucosa. La activación de monosacáridos con UPT es un
mecanismo habitual en las células. Esta activación determina que el
monosacárido sea aprovechado para la formación de ósidos, ya que la formación
del enlace o-glucosidico es un proceso endergónico que necesita energía
aportada por la hidrolisis del UDP del monosacáridos
Para la síntesis de glucógeno a partir de
moléculas de UDP-glucosa, se necesitan:
Ÿ Una
molécula preexistente del glucógeno o un cebador como la glucogenina.
Ÿ La
enzima glucógeno sintasa, cuyo papel será alargar las cadenas lineales del
glucógeno mediante la adicción de
moléculas de glucosa precedentes de la UDP-glucosa, uniéndolas mediante la
adición de moléculas de glucosa procedentes de la UDP-glucosa, uniéndolas
mediante enlaces o-glucosídico (α 1 → 4) con la cadena preexistente del
glucógeno.
Ÿ La
enzima ramificante, cuyo papel es crear los puntos de ramificación mediante
enlaces (α 1 → 6). Esta enzima tiene una actividad glucosil transferasa, que
transfiere parte de la cadena de moléculas de glucosa en enlaces α 1 → 4, uniéndola
al glucógeno a través de un enlace α 1 → 6. (Feduchi Elena, 2010)
CONCLUSIÓN
Finalizando nuestro informe podemos
destacar la importancia de incluir a los hidratos de carbono en nuestra dieta
humana. Los hidratos de carbono son los componentes de la dieta que menos
cantidad de calorías aportan por unidad de peso –alrededor de 4 Kcal. /g–.
Mientras que esta relación en el alcohol es de 7 Kcal. /g. Los carbohidratos
deberían representar el 50% de nuestro aporte energético diario, y las grasas
no deberían superar el 30. Los carbohidratos (hidratos de carbono) nos engordan
con más facilidad
Cuando los niveles de glucosa en la sangre aumentan, el páncreas estimula la producción
de la hormona insulina, la
cual se encarga de guardar la glucosa en las despensas. Sin embargo, ésta no es
su única misión: a la vez que guarda, también se asegura que la grasa guardada
no se queme. Por esta razón, y aunque la insulina es muy importante para mantenernos vivos, hay que
evitar producir grandes cantidades. Sin duda, la forma más efectiva de
controlar los niveles de insulina es a través de comer alimentos con un bajo
índice glicémico.
Recordemos que a mayor índice glicémico, mayor producción de insulina. Además, los primeros tienen un efecto más saciante que las grasas
Aquellas personas que desean
adelgazar, deberían procurar sustituir el consumo de grasas por hidratos de
carbono, lo que supondría una mejora en su dieta y permite perder peso a largo
plazo.
La mayoría de las personas
identificamos los hidratos de carbono con los alimentos ricos en energía y
quienes se preocupan por su peso, además procuran esquivarlos a la hora de
diseñar su dieta. Las verduras, las carnes y los pescados son los alimentos más
indicados para evitar los carbohidratos.
Tan importante como incluir en
nuestra dieta carbohidratos es tomar proteínas, presentes en carne, pescado,
huevos y frutos secos. La proporción de alimentos ricos en carbohidratos, que
se recomienda comer, debe ser el doble que la correspondiente a alimentos con
proteínas.
Bibliografía
• Mathews van Holde.
Bioquímica, E. M.–I. (2015). metabolismo hidrato de carbono. Editorial Omega.
Ediciones varias.
Bioquímica, P. (.
(2015). Metabolismo de hidratos de carbono. PORTOVIEJO: Editorial
Limusa.
Bioquímica., P. (.
(2015). Portoviejo : Editorial Limusa.
Feduchi Elena, B. I.
(2010). Bioquímica. Conceptos esenciales. Madrid: Ed. Médica
Panamericana.
Feduchi, B. R. (2015). metabolismoo
de hidrato de carbono . Bioquímica Conceptos.